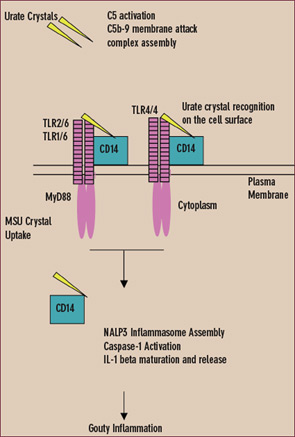
Experimental gouty inflammation is triggered by IL-1 receptor signaling by resident cells at the site of urate crystal injection.13 Macrophage recognition and uptake of urate crystals by the cell surface Toll-like receptors (TLRs) 2 and 4 and by signaling of the shared intracellular TLR adaptor protein MyD88 drives both pro- and anti-inflamamtory cytokine expression.14 Depending on the locus of experimental urate crystal injection, TLR2 and TLR4 expression determines dominant anti-inflammatory effects (such as induction of transforming growth factor b) or pro-inflammatory effects in vivo. However, MyD88, which also is a critical adaptor protein for the IL-1 receptor, is central to both urate crystal uptake by cultured macrophages and urate crystal-induced inflammation in vivo.13,14
In the cytoplasm, urate crystals also employ the cytoplasmic pattern recognition receptor NALP3 (cryopyrin) to drive inflammation.11,12 Upon ligand recognition, NALP3, like other members of the nucleotide-binding oligomerization domain (NOD)–like receptor family, assembles with other proteins in large complexes termed inflammasomes. When NALP3 complexes with caspase-recruiting domain (CARD)–carrying adaptor proteins, activation of the latent inflammatory mediator caspase-1 is stimulated. The activated NALP3 inflammasome complex thereby induces proteolysis of pro–IL-1b and subsequent maturation and release of IL-1b.11


