(click for larger image)
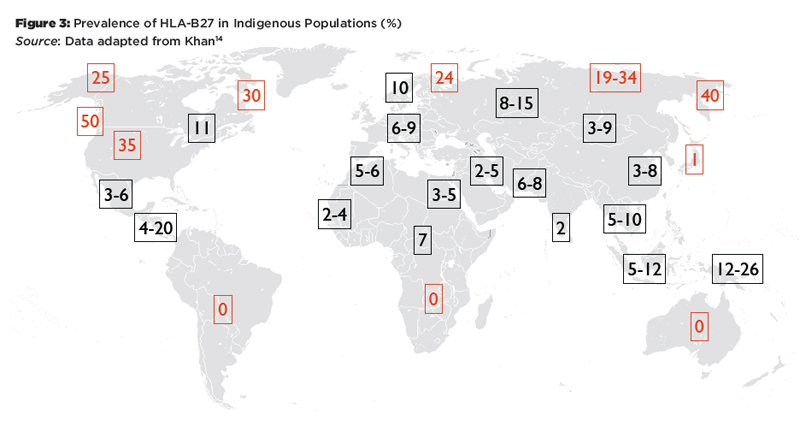
Figure 3: Prevalence of HLA-B27 in Indigenous Populations (%)
Source: Data adapted from Khan14
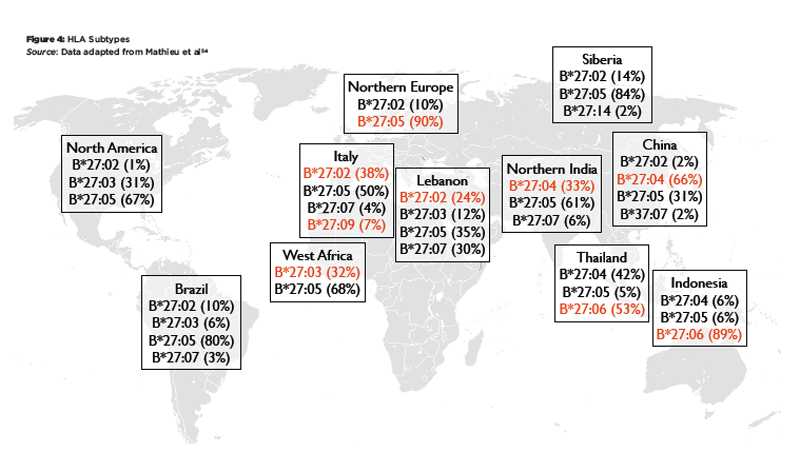
These patterns reflect two major influences that have shaped MHC profiles in populations: migration and natural selection. For example, HLA-B27 is absent in the indigenous populations of South America, and the HLA-B27 alleles prevalent on this continent today represent genetic admixture from Europe and Africa.15 It has been argued that the lack of HLA-B27 in South America and Southern Africa and the high frequency of HLA-B*27:06 in Southeast Asia may be the result of selective pressure by malaria or other infectious diseases.16 On the other hand, there is evidence that HLA-B27 is associated with HIV elite controller status and higher virus clearance rates after acute HCV infection.17 Thus, being HLA-B27 positive may be a selective advantage in certain circumstances, which might explain the high prevalence in circumpolar regions.
(click for larger image)
Figure 4: HLA Subtypes
Source: Data adapted from Mathieu et al54
The 2009 NHANES (National Health and Nutrition Examination Survey) study examined the prevalence of axial SpA and HLA-B27 in subjects representative of the U.S. population. Interestingly, HLA-B27 was significantly less frequent in the older age brackets (3.6% in 50–69-year-olds vs. 7.3% in 20–49-year-olds).18 This difference remained statistically significant after adjusting for sex and race/ethnicity, suggesting that HLA-B27 positivity might be associated with early mortality (e.g., via effects on atherosclerosis or cancer), a surprising finding that needs to be confirmed in an independent cohort.