Moreover, some of these cells expressed T resident memory (TRM) cell markers. The TRM cells are sentinels in peripheral tissues with traditional α,β T cell receptors that respond rapidly to pathogens.24,25 The cells can also promote inflammation and have been implicated in the pathogenesis of several diseases, including psoriasis, mycosis fungoides and juvenile inflammatory arthritis.26-28
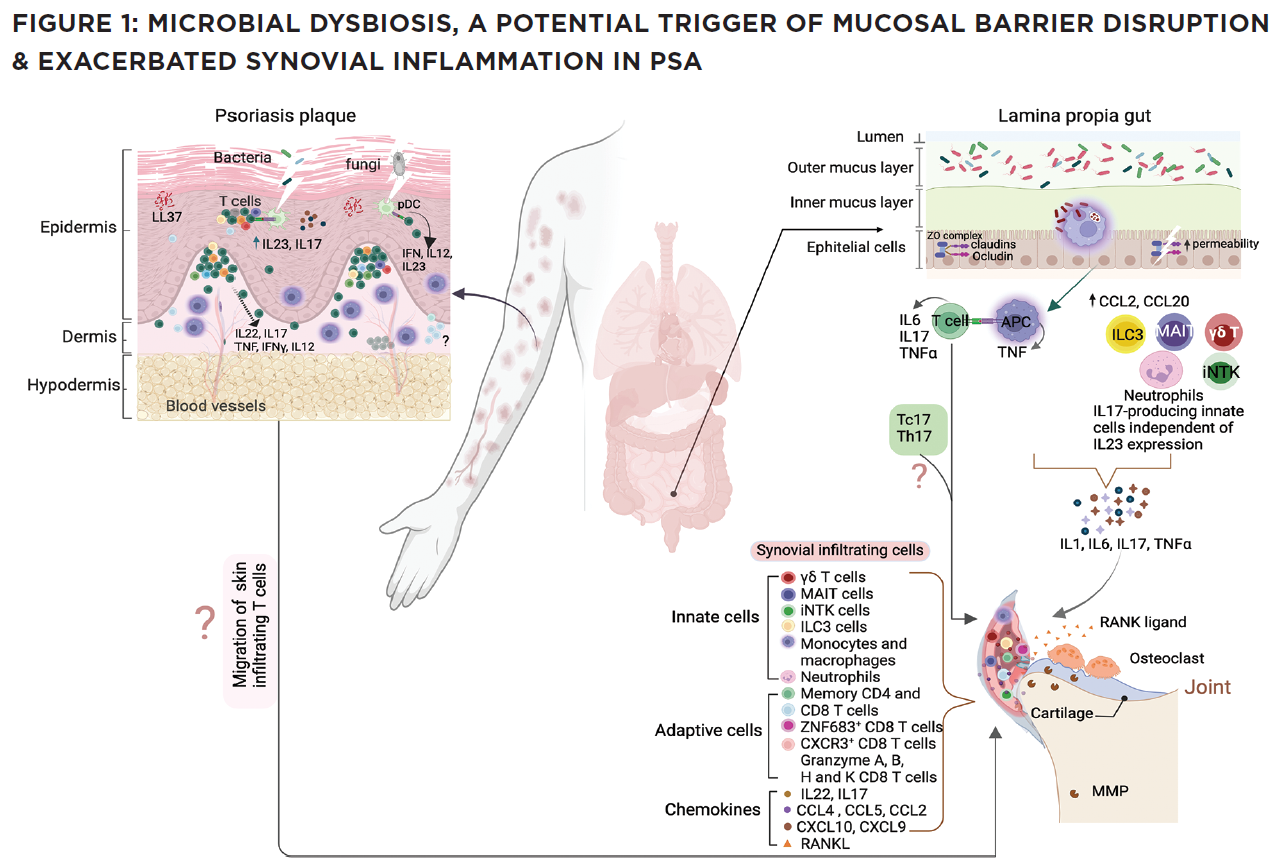
(CLICK TO ENLARGE) In this model, impaired pathogen immunity in PsA facilitates colonization of the skin and gut with bacteria that disrupt barrier integrity. Microbial products stimulate the release of chemokines (CCL2, CCL20), which draw innate immune cells including monocytes, macrophages, neutrophils, gamma delta T cells, innate lymphoid cells 3 (ILC), mucosal-associated invariant T cells (MAIT) and invariant natural killer T (iNKT) cells into target tissues. Innate and acquired immune cells secrete IL-17 and IL-23, inducing the proliferation of keratinocytes in the skin and the production of proinflammatory cytokines (IL-1, IL-6, TNF, IL-17) by resident synovial stromal cells. These cytokines trigger the secretion of additional chemokines responsible for the attraction and differentiation of CD4 and CD8 T cells in the skin and the joint. Tissue-infiltrating CD4 and CD8 T cells differentiate into Th17 or Tc17 cells, which release IL-17, a cytokine responsible for the generation of chemokines that attract neutrophils (CXCL1, CXCL2) and coordinate the migration of CXCR3+ T cells (CXCL9 and CXCL10). The presence of cycling, activated ZNF683+ resident memory CD4 and CD8 T cells in the synovia and skin of PsA patients release an array of cytokines. Expression of CD49a on the surface of synovial CD8 T cells indicates their potential bidirectional migration from or to the skin or the intestine. In summary, although Th17 cells are currently considered the primary drivers of PsA pathogenesis, novel findings support the participation of innate immune cells as a source of IL-17 and unexpectedly reveal the potential of resident memory CD8 T cells to mediate tissue inflammation and damage in PsA patients.