Age-Related Change
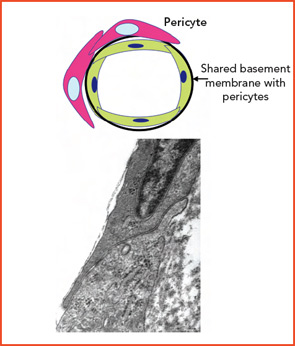
The aging process is associated with fragmentation of elastic fibers, depletion of elastica, and an increase in collagen deposition. The most significant changes occur in conjunction with an increase in vessel-wall concentrations of matrix metalloproteinases in muscular/elastic vessels. Coupling of vessel and tissue proteins with reducing sugars leads to production of advanced glycation end products that alter the function of normal proteins and may change their antigenicity. Evidence of progressive EC dysfunction includes a reduced capacity to produce nitric oxide and an increased engagement of pathways of oxidative stress that enhance risk of atherosclerosis and thrombosis. These changes may vary in severity in different sites.8-10
Over our lifespans, we experience innumerable interactions with our environment, acquiring, in some cases, inanimate materials by transcutaneous, airborne, or ingested pathways. Other acquisitions include living materials that were transmitted as early as during gestation (e.g., microchimerisms, or cells from our mothers), or later from others or the environment (e.g., infectious agents, especially herpes viruses). Each may have favorite locations within tissue, including vessels, where they may be of no consequence or could lead to overt infection or changes in tissue antigen expression.